Физиология движений. Все наши движения могут быть подразделены на произвольные, активные, совершаемые по собственному желанию; на пассивные, совершаемые под влиянием воздействия внешней силы; рефлекторные, осуществляющиеся на уровне сегментарного аппарата, и на ряд непроизвольных или насильственных движений, возникающих при патологических процессах в различных отделах нервной системы, о чем будет идти речь в соответствующих главах. Выполнению любого произвольного действия предшествует построение плана этого действия, создается его модель. Реализация модели, т. е. выполнение соответствующего действия, невозможна при помощи одних лишь эфферентных систем, посылкой эфферентных импульсов к исполнительным аппаратам. Суставно-мышечный аппарат человека имеет такое большое количество степеней свободы, что выполнение любого целенаправленного движения без афферентного контроля невозможно. В процессе выполнения движения в центральную нервную систему поступают афферентные импульсы, сигнализирующие о положении движущейся конечности в пространстве. Поступающая информация сравнивается с созданной ранее моделью, движение постоянно корригируется дополнительными афферентными импульсами, чем и обеспечивается точность его исполнения. И в самом деле, трудно ожидать от человека успешного движения рукой, если не поступили сигналы о том, в каком положении она находится в данный момент. Действие проприоцептивной афферентной системы к тому же дополняется действиями других систем — зрительной, вестибулярной, слуховой и др. Таким образом, произвольное движение обусловлено сложными взаимодействиями ряда различных физиологических систем.
Движения характеризуются не только определенной силой, но и дифференцированностью, четкостью, быстротой и ловкостью, слаженностью работы мышечных групп (при постоянном соответствии функции кровеносных сосудов, питающих мышцы, и лимфатических путей).
Как было сказано в разделе нейрофизиологии, наши произвольные движения, помимо обусловленности рефлекторными механизмами, подчинены мотиву действия, осуществлению тех или иных, жизненных потребностей.
Весь сложный комплекс реакций от периферического рецептора к мозговой коре и далее через кору и спинной мозг к мышце, как и все наши рефлексы, совершается очень быстро. Ведь трудно отделить наш приказ к действию от выполнения этого действия, а предуготовленность всех мышц даже предшествует самому действию.
Произвольные целенаправленные движения не даны от рождения в законченном виде уже по одному тому, что мозг человека продолжает развиваться после рождения. Человек обучается этим движениям постепенно с раннего детства, закрепляя их через огромное количество повторений в динамические стереотипы, образы движений. Чтобы убедиться в сказанном, достаточно понаблюдать за развитием двигательных навыков у ребенка. Сколько повторений необходимо, чтобы научиться пользоваться ложкой и вилкой. Сколько усилий должна затратить, сколько настойчивости должна проявить маленькая девочка для того, чтобы научиться стоять на одной ножке, прыгать сначала через неподвижную, а затем вертящуюся веревку. Что же говорить об игре на рояле или другом музыкальном инструменте?
Совершенно ясно, что связь в двигательных системах устанавливается постепенно. Если эта связь «потом постоянно в течение всей жизни расширяется и совершенствуется, естественно предположить, что и первое время индивидуального существования высших животных и особенно человека, когда последний месяцами обучается управлять своими первыми движениями, идет именно на образование этой связи».
Произвольные движения осуществляются через сегментарно-рефлекторный аппарат спинного мозга и ствола мозга, а также через систему нейронов, которые по отношению к сегментарно-рефлекторному аппарату могут быть названы центральными нейронами.
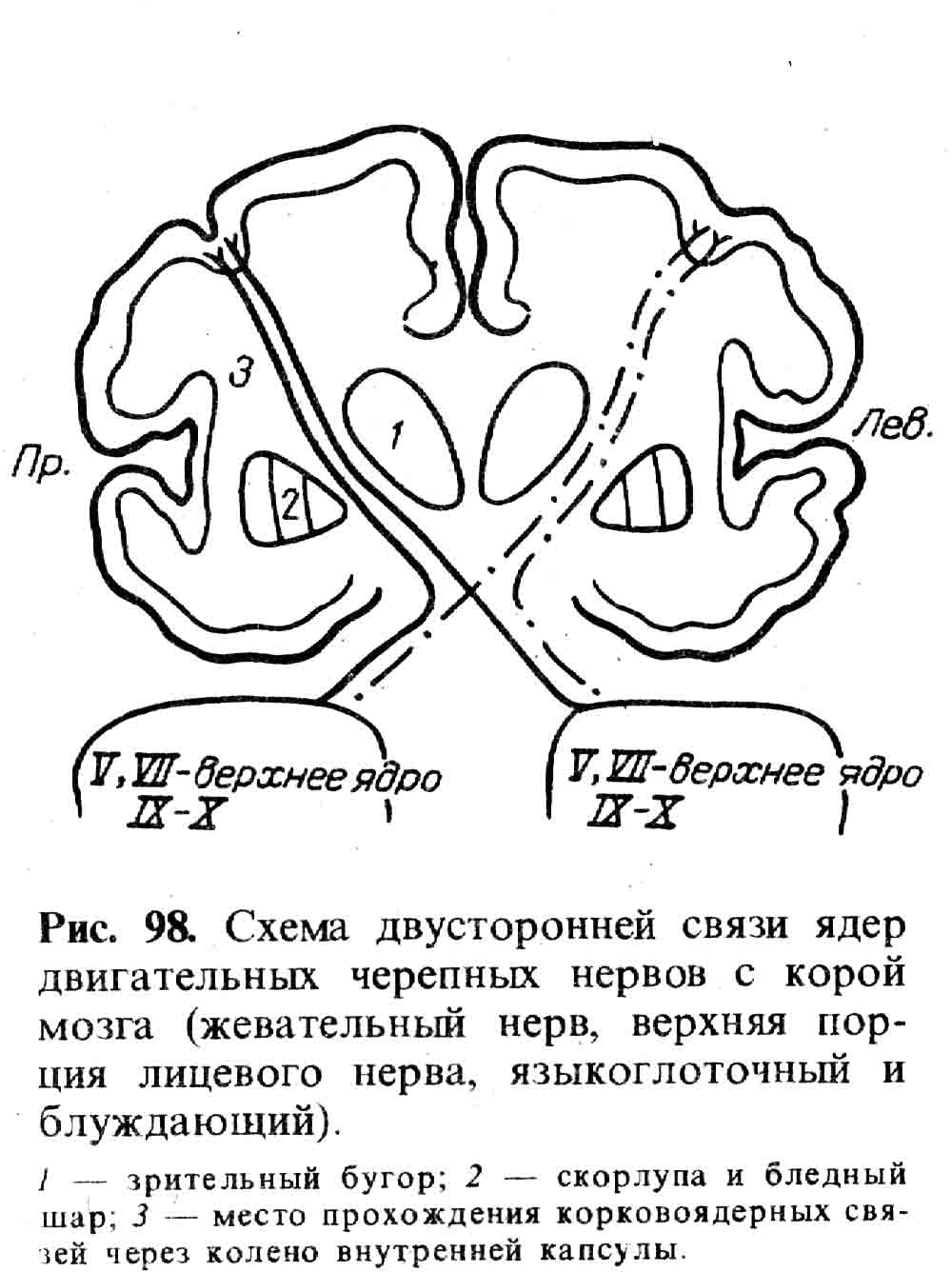
Анатомия. Основной двигательный путь, через который осуществляется связь моторной коры (см. рис. 20) с мышцами, состоит из двух нейронов: центрального, идущего от коры к спинному мозгу, и периферического двигательного нейрона, берущего начало от клеток передних рогов спинного мозга, аксон которых в составе периферического нерва и достигает мышц (см. рис. 24). Совокупность центральных нейронов образует так называемый пирамидный путь. Началом его являются преимущественно крупные пирамидные нейроны— клетки Беца (см. рис. 100), расположенные в V слое коры передней центральной извилины и парацентральной дольки и ряда других (менее значимых) областей коры. 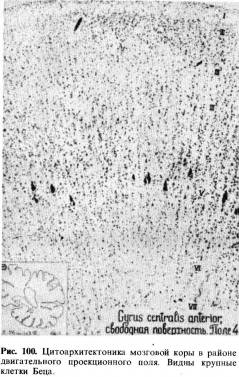
Аксоны клеток, образующих пирамидный путь, направляясь вниз, располагаются под корой в виде лучистого венца, а дальше, постепенно сближаясь друг с другом, занимают место между серыми подкорковыми ядрами, образуя заднее бедро и колено внутренней капсулы (capsula interna). Волокна, идущие через колено, носят название кортико-нуклеарных, или кортико-ядерных волокон, так как они заканчиваются у ядер двигательных черепных нервов.
Несколько слов о той терминологии, которая употребляется при описании тех или иных путей. Названия, их слагаются из названия того отдела, где расположены клеточные тела нейронов, откуда начинается их путь, и из названия места окончания пути, т. е. места расположения тех клеток, к которым подходят аксоны данных нейронов. Так, кортико-спинальный путь идет от коры к спинному мозгу, кортико-нуклеарный — от коры к ядрам черепных нервов; вестибуло-спинальный путь идет от вестибулярного ядра продолговатого мозга к спинному мозгу и т. д.
Волокна пирамидного пути, пройдя через заднее бедро внутренней капсулы, попадают в ножку мозга (через ножку мозга проходят как кортико-спинальные, так и кортико-нуклеарные волокна), далее в мост и продолговатый мозг, всюду располагаясь вентрально (рис. 33). 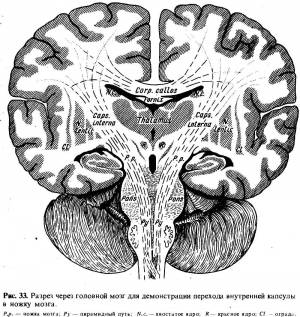
В срезе продолговатого мозга эти волокна занимают участок, напоминающий по форме пирамиду, что и послужило основанием назвать их пирамидным путем. В нижних отделах продолговатого мозга почти все кортико-спинальные волокна совершают переход на противоположную сторону спинного мозга, размещаясь в боковых его столбах и, постепенно убывая, заканчиваются в клетках передних рогов всех сегментов спинного мозга. Волокна, не совершающие перехода на противоположную сторону (таких примерно 15%), идут в передних столбах спинного мозга (рис. 34), также заканчиваясь в клетках передних рогов. Неперекрещивающиеся волокна служат в основном для передачи импульсов к мускулатуре, расширяющей грудную клетку при акте дыхания. Эти мышцы как действующие одновременно с обеих сторон получают импульсы и по перекрещенным, и по неперекрещенным путям. Не только дыхательная мускулатура, но и мышцы верхней части лица, жевательные, мышцы мягкого неба, голосовых связок, промежности — все они как действующие всегда одновременно с обеих сторон связаны через кортико-нуклеарные или кортико-спинальные волокна с обоими полушариями головного мозга (см. рис. 98). 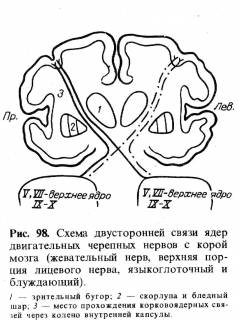
Мышцы нижней половины лица, мышцы языка, руки, ноги, туловища (кроме дыхательных) получают импульсы только от противоположного полушария головного мозга.
Периферический двигательный нейрон связан с мышцами соименной стороны. Таким образом, весь двухнейронный двигательный путь, например, для мышц левой руки идет следующим образом: клетки нижних отделов правой передней центральной извилины, заднее бедро правой внутренней капсулы, правая ножка мозга, правая половина моста и продолговатого мозга, перекрест на границе продолговатого и спинного мозга, далее левый боковой столб спинного мозга, клетки передних рогов шейного отдела левой половины спинного мозга. Аксоны этих клеток образуют передние корешки, которые участвуют в образовании левого плечевого сплетения и периферических нервов, которые подходят к мышцам левой руки.
Информация предоставлена medvuz.info
Похожие статьи
Добавь в закладки
|