В понятие
«сегментарно-рефлекторный аппарат спинного мозга» входит его серое вещество с
передними и задними корешками, из которых каждый связан с определенным участком
мышечной системы (передний, эфферентный, корешок) и с определенным участком
кожи (задний, афферентный, корешок). Этим определяется название «сегментарный
аппарат». Но это и отдел переключения безусловных рефлексов, примером чего
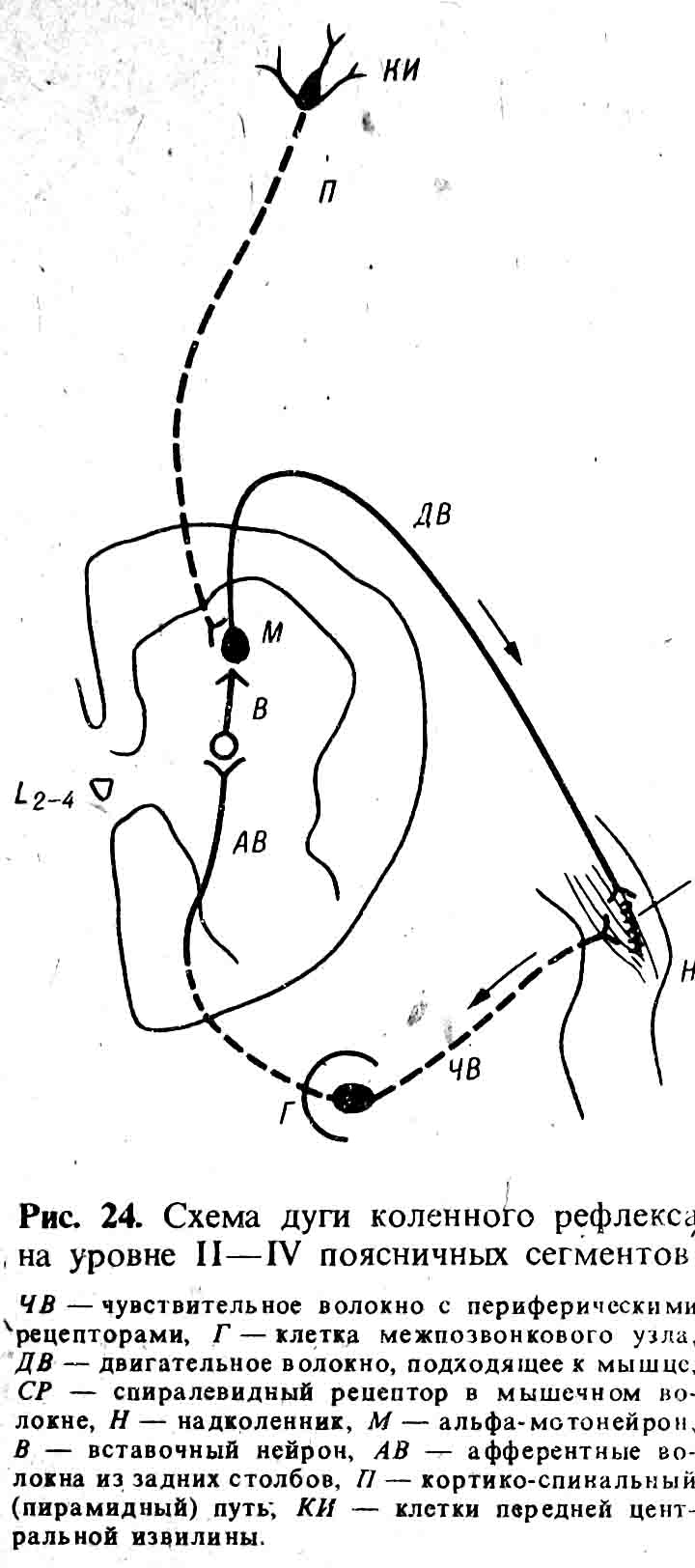
может служить хотя бы колейный рефлекс (рис. 24). 
Этим собственно определяется
название «рефлекторный аппарат». Нужно запомнить, что афферентной частью в дуге
сухожильных рефлексов (см. ниже) являются коллатеральные волокна от задних
столбов к передним рогам. Помимо сегментарно-рефлекторной части, в спинном
мозге имеются проводники, как имеются они и во всех других этажах нервной
системы, поскольку все отделы головного и спинного мозга связаны друг с другом.
Всякий спинномозговой рефлекс
осуществляется не одним, а несколькими сегментами спинного мозга, однако один
из сегментов всегда имеет преимущественное значение. Межсегментные связи
обеспечивают координированную деятельность сегментарного аппарата спинного
мозга.
Спинальный сегментный
аппарат обеспечивает постоянное напряжение скелетных мышц, сопротивление их
растяжению, что носит название мышечного тонуса. Мышечный тонус, отражающий
степень деятельности состояния периферического двигательного нейрона,
называется контрактильным, проприоцептивным рефлекторным тонусом.
Регуляция мышечного тонуса
осуществляется довольно сложными механизмами. Имеется два вида различных по
величине мотонейронов — «фазические» мотонейроны (α-мотонейроны большие),
которые иннервируют так называемые белые мышечные волокна, способные совершать
быстрые сокращения, и «тонические» мотонейроны (α-мотонейроны малые), которые
иннервируют красные мышечные волокна, играющие важную роль в поддержании тонуса
и позы. Кроме α-мотонейронов, имеются γ-мотонейроны, которые составляют около
трети общего числа клеток передних рогов (по Р. Граниту). Аксоны α- и
γ-мотонейронов идут на периферию в составе передних корешков и периферических
нервов. Аксоны α-мотонейронов заканчиваются концевыми пластинками на мышечных
волокнах (экстрафузальное мышечное волокно). Аксон же γ-нейрона подходят к
обоим концам так называемых интрафузальных мышечных волокон. В средней
(экваториальной) части этих волокон помещается рецептор афферентного волокна,
имеющий вид спирали. Импульс γ-нейрона вызывает сокращение мышечных элементов
интрафузального волокна. Это приводит к растяжению его экваториальной области и
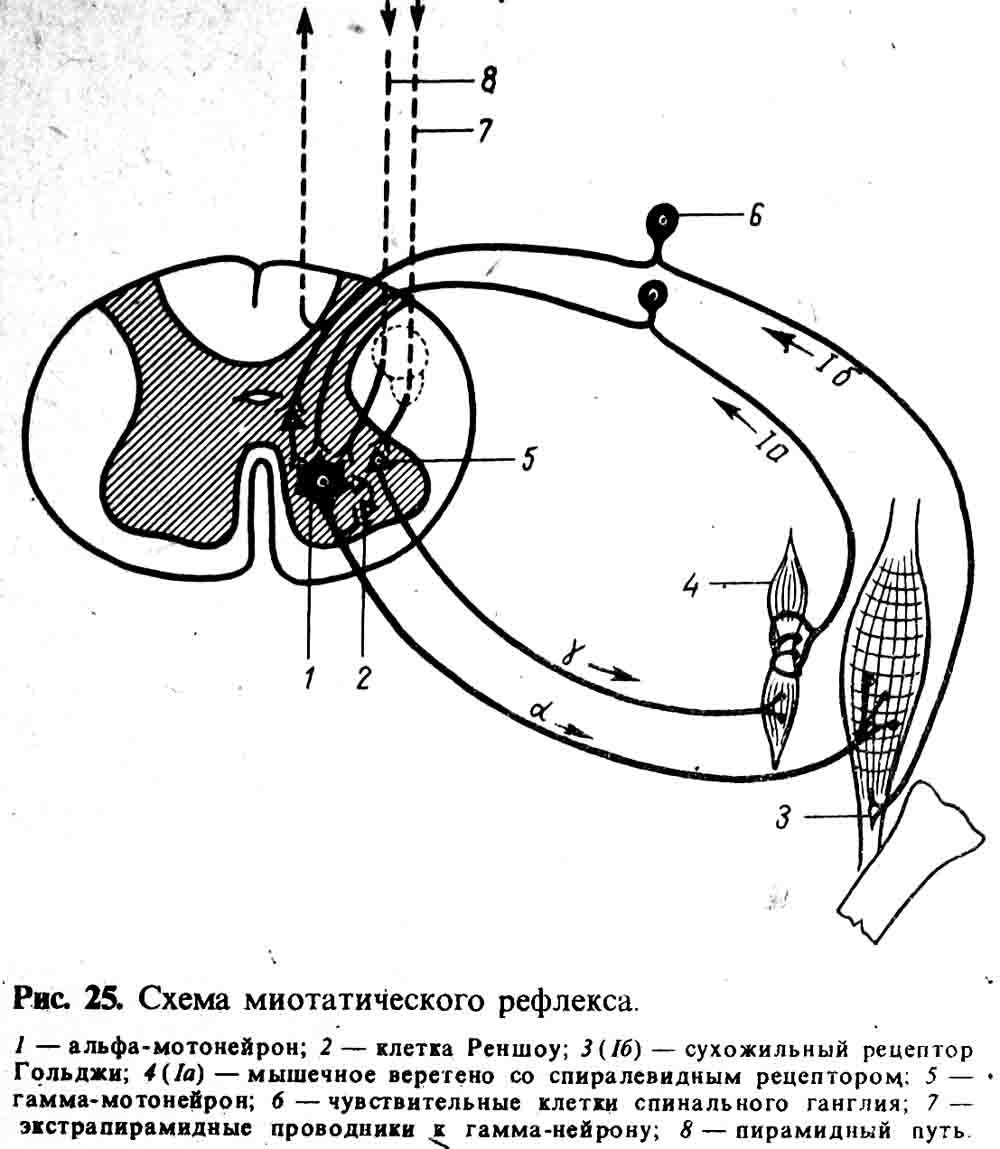
возникновению деполяризации в расположенной здесь спирали рецептора волокна 1а
(рис. 25). 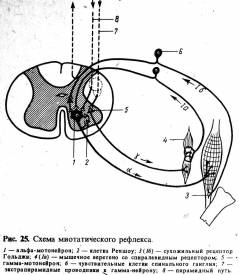
Возникающее в результате этого возбуждение по афферентному волокну
переносится на α-мотонейрон и вызывает тоническое напряжение мышцы
По γ-эфферентным волокнам
импульсы поступают непрерывно, поэтому интрафузальные волокна всегда находятся
в состоянии некоторого сокращения, что в свою очередь влечет за собой
непрерывность импульсации от интрафузальных волокон, адресуемой к
α-мотонейронам спинного мозга.
Описанный рефлекс
растяжения регулируется центральными (супрасегментарными) приводами к α- и
γ-мотонейронам спинного мозга. Такие приводы поступают от коры полушарий
большого мозга, стриопаллидарной системы, зрительного бугра, среднего мозга,
продолговатого мозга и мозжечка (см. рис. 25).
Часть дендритов нервных
клеток спинальных ганглиев (волокна 1б, рис. 25) заканчиваются не в мышечных
веретенах, а в особых рецепторах сухожилий (сухожильные рецепторы Гольджи).
Усилие, создаваемое напрягающейся мышцей, вызывает возбуждение рецептора
Гольджи, по аксону которого (через вставочную клетку) идет торможение
активности α-мотонейрона, если она является излишней. Включение в действие
антагонистов предохраняет мышцу от перенапряжения. Саморегулирующий механизм
тонуса мышц еще связан и с клетками Реншоу, которые оказывают тормозящее
действие на α-мотонейрон. Все описанное демонстрирует сложность механизма,
регулирующего мышечный тонус и готовность к строгому соответствию сокращения
мышц текущим потребностям.
О сложности обеспечения
мышечного тонуса, этого важного фактора движения, можно судить по
представленной упрощенной схеме регуляции его на уровне спинального сегмента,
представляющей уже не дугу, а замкнутое кольцо. Вставочные тормозные и
активирующие импульсы из подкорковых образований, поступающие (через
ретикулоспинальный путь) на α-нейрон, вставочные клетки Реншоу и
пресинаптическое торможение — все это тонкие механизмы саморегулирующего
прибора. Патологическое повышение тонуса мешает активным и пассивным движениям,
и резкое повышение его вызывает длительное пребывание суставов в приданном
положении.
Согласованное напряжение и
расслабление мышц агонистов и антагонистов обеспечиваются межсегментарными
(спинальным и стволовым) механизмами. Они выражаются в наличии реципрокной
(обоюдной, взаимной) иннервации мышц, обеспечивающих движение, и сопряженного
сокращения мышц для фиксации сустава (рис. 26). 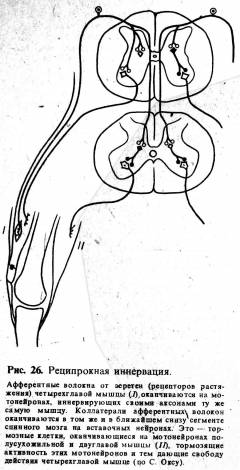
Так, например, при сгибательном
движении одновременно с напряжением сгибателей происходит реципрокное
расслабление тонуса антагонистов — разгибателей и, наоборот, при разгибании —
реципрокное расслабление сгибателей. При выполнении мелких движений кистью
одновременно сама собой фиксируется рука в локтевом и плечевом суставах путем
сопряженного сокращения соответствующих мышц. Можно привести множество подобных
примеров, ибо в каждом движении имеются эти механизмы.
Каждому произвольному
двигательному акту предшествуют позиционное возбуждение и установка мышц,
дающая возможность совершить движение четко и без ущерба для организма.
Например, подниманию левой ноги предшествует напряжение мышц правой нош и
туловища для удержания равновесия. Это все срабатывается автоматически, без
участия сознания, однако лишь до тех пор гладко и безупречно, пока сохранены
все,нервные связи, афферентные и эфферентные нейроны.
В регулировании мышечного
тонуса в клинике учитывается немаловажная роль, принадлежащая нервным
импульсам, поступающим по кортико-спинальному (пирамидному) пути на
сегментарно-рефлекторный аппарат, именно на эфферентные его отделы (на клетки
передних рогов), которые обеспечивают произвольные движения. Эти импульсы,
постоянно поступающие на эфферентные приборы (как ответ на массу раздражений,
идущих в мозг из собственных тканей и из внешнего мира), осуществляя
саморегуляторный механизм, притормаживают, регулируют нервные импульсы, которые
идут по рефлекторным дугам (кольцам) и обеспечивают, как показано выше, мышечный
тонус в выгодных для организма пределах. В тех случаях, когда импульсы по
пирамидному пути не поступают на сегментарно-рефлекторный аппарат,
несдерживаемый поток раздражений по спинальным рефлекторным кольцам поступает в
мышцы, тем самым вызывая избыточное (невыгодное) повышение мышечного тонуса.
Тот же принцип сегментарно-рефлекторного
построения лежит в основе функции ствола мозга, где роль корешков играют
черепные нервы, а аналогами передних рогов являются двигательные ядра этих
нервов. Черепные нервы участвуют в образовании рефлекторных дуг многообразных
безусловных рефлексов, например глоточного, небного. Богатые межсегментарные
связи обеспечивают механизмы и более или менее сложных сочетанных движений,
куда относятся акт глотания, рвотный рефлекс, жевание, чиханье, мигание,
голосообразование, дыхание, сердечная деятельность, слезотечение и
слюноотделение. Координация функций различных систем, участвующих в
перечисленных автоматических актах, происходит за счет сетевидного образования
продолговатого мозга и моста, которое связывает ядра черепных нервов не только
между собой, но и с клетками верхних отделов спинного мозга. Корково-ядерные,
или кортико-нуклеарные, связи обеспечивают регуляцию мышечных сокращений и
напряжений в соответствующих отделах.
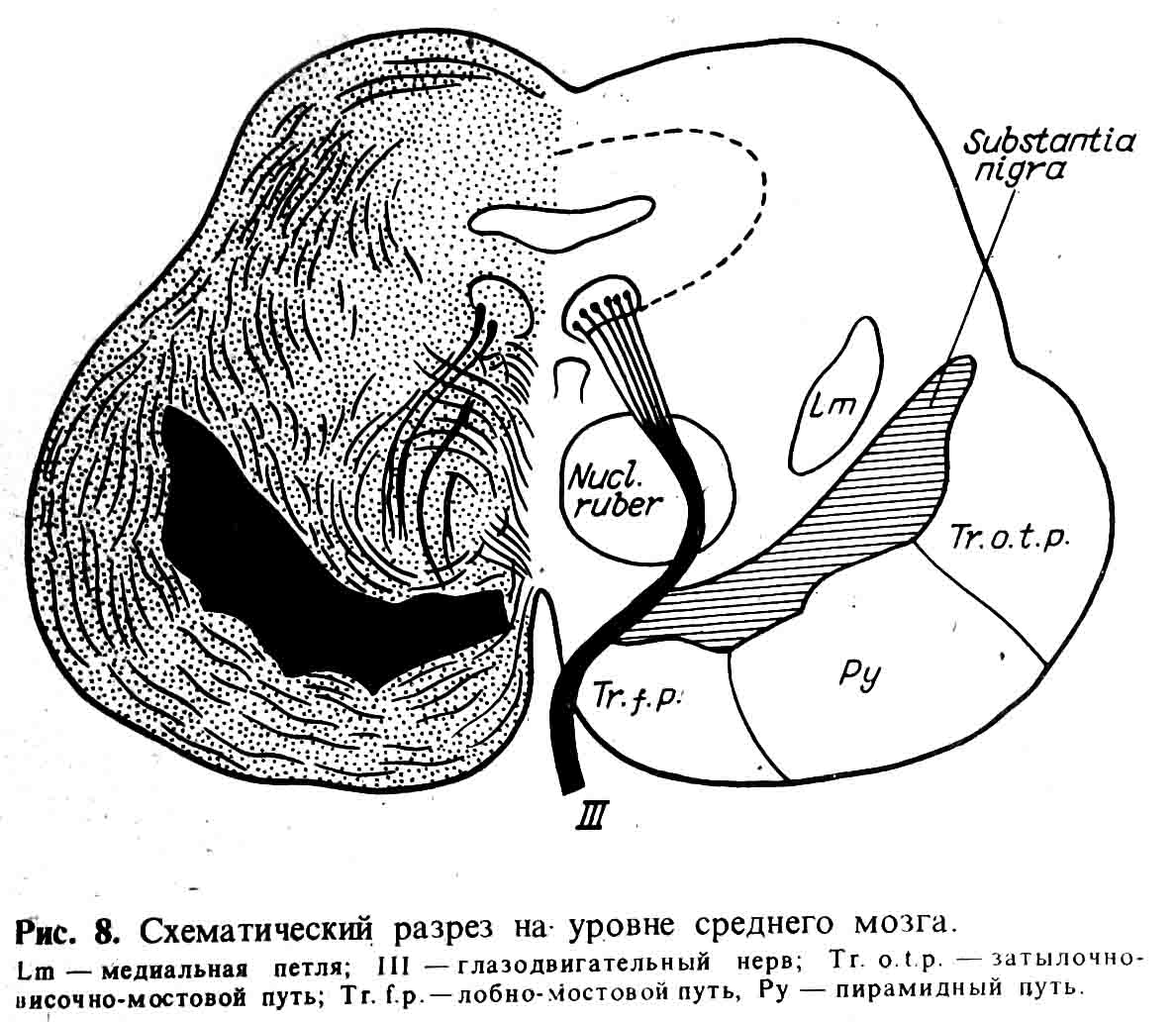
Еще более сложной частью
сегментарно-рефлекторного aппaрата является средний мозг, т. е. ножки мозга и
четверохолмие; (см. рис. 8) с двумя парами ядер черепных нервов — III и IV. 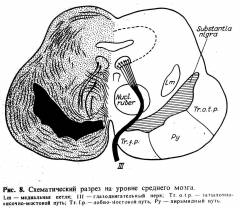
Как
это показал еще Е. К. Сепп, четверохолмие является отделом, обеспечивающим быструю
ориентировку и установку головы и тела человека при световых и звуковых
раздражениях, а красные ядра и ретикулярная формация играют большую роль в
регуляции мышечного тонуса. Они получают афферентные импульсы из полушарий
мозжечка, от коры головного мозга, зрительного бугра и полосатого тела.
Эфферентные импульсы отсюда идут как в кору головного мозга, зрительные бугры,
полосатые тела, так и в спинной мозг.
В эксперименте на животных
Ч. С. Шеррингтоном (Ch. Sherrington) показано, что при перерезке ствола мозга
между верхним и нижним четверохолмием позади красных ядер и, значит, при
отделении ствола от полушарий мозга, особенно, при одновременном повреждении
красных ядер, наступает децеребрационная ригидность, т. е. резкое усиление
проприоцептивного рефлекторного тонуса с преобладанием его в разгибательной
группе мышц. Децеребрированное животное стоит, но стояние его неустойчивое,
пассивное: от малейшего толчка животное падает и не может само подняться.
В клинике явления типа
децеребрационной ригидности могут наблюдаться при травмах и кровоизлиянии в
мозговой ствол, а также при других патологических процессах. Информация предоставлена medvuz.info
Похожие статьи
Добавь в закладки
|