Мозг
1.ДВИГАТЕЛЬНЫЙ АППАРАТ ГЛАЗ
1.1.Морфо-физиологические особенности глазодвигательного аппарата
1.2.Бинокулярная координация движений глаз
1.3.Временные характеристики движений глаз
2.ЦЕНТРАЛЬНАЯ ОРГАНИЗАЦИЯ ДВИЖЕНИЙ ГЛАЗ
2.1.Первый уровень глазодвигательной системы
2.2.Второй уровень глазодвигательной системы
2.3.Третий уровень глазодвигательной системы
2.4.Четвертый уровень глазодвигательной системы
3.ФИЛОГЕНЕТИЧЕСКИЕ АСПЕКТЫ ГЛАЗОДВИГАТЕЛЬНОЙ АКТИВНОСТИ
4.ИЕРАРХИЧЕСКОЕ УПРАВЛЕНИЕ ДВИЖЕНИЯМИ ГЛАЗ
Проблема взаимосвязи движений глаз с перцептивными функциями зрения отчетливо прозвучала в работах И.М. Сеченова еще в прошлом столетии. В последующем развитии концепций восприятия эта проблема вызывала очень много споров и сохранила свою дискуссионность до настоящего времени. При этом обсуждение центрального вопроса о так называемой "построительной" функции движений так и не привело исследователей к единому мнению.
По сей день сторонники моторной концепции отводят движениям глаз ведущую роль в формировании зрительного образа. Приверженцы сенсорной парадигмы, напротив, считают, что перцептивные функции, как таковые, очень мало зависят от моторных компонентов зрения. Обе точки зрения развернуто аргументируются экспериментальными данными и концептуальными моделями. Глобальная проблема взаимодействия сенсорных и моторных компонентов зрения коррелирует с рядом более частных вопросов, от решения которых зависит развернутый ответ на центральный вопрос. Как соотносится моторика глаз с другими видами мышечной активности? Какими механизмами (нервно-мышечными и нейронными) определяется жесткая дихотомия глазных движений на саккадические и плавные? В какой последовательности развиваются разные формы глазодвигательной активности в филогенезе и онтогенезе? В чем состоит вклад разных мозговых уровней в управление движениями глаз? Как влияют прямые и обратные нервные связи на организацию саккады? В какой мере специфика глазных движений коррелирует со структурой поля зрения?
1. ДВИГАТЕЛЬНЫЙ АППАРАТ ГЛАЗ.
1.1.МОРФО-ФУНКЦИОНАЛЬНЫЕ ОСОБЕННОСТИ ГЛАЗОДВИГАТЕЛЬНОГО АППАРАТА
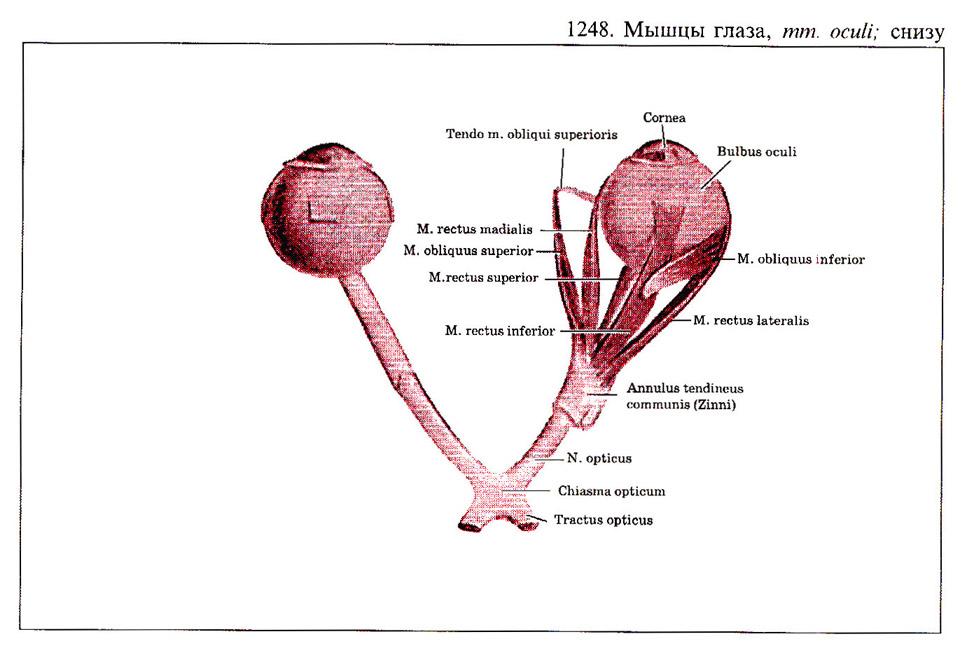
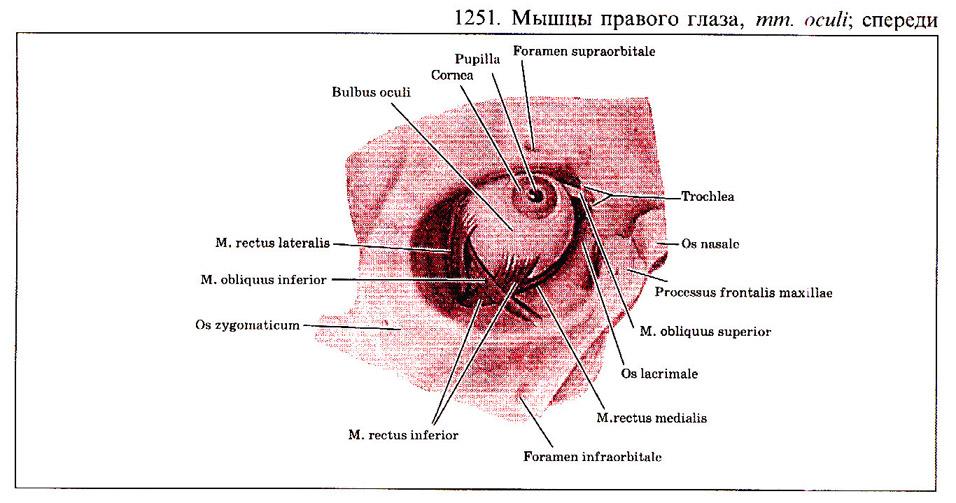
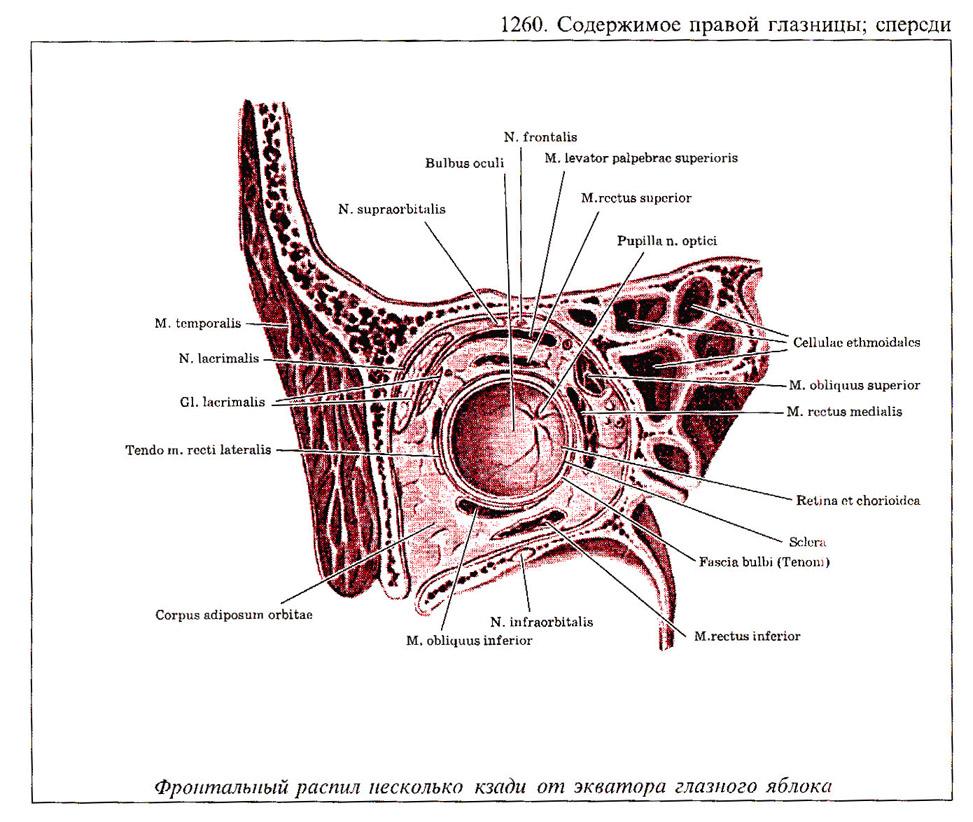
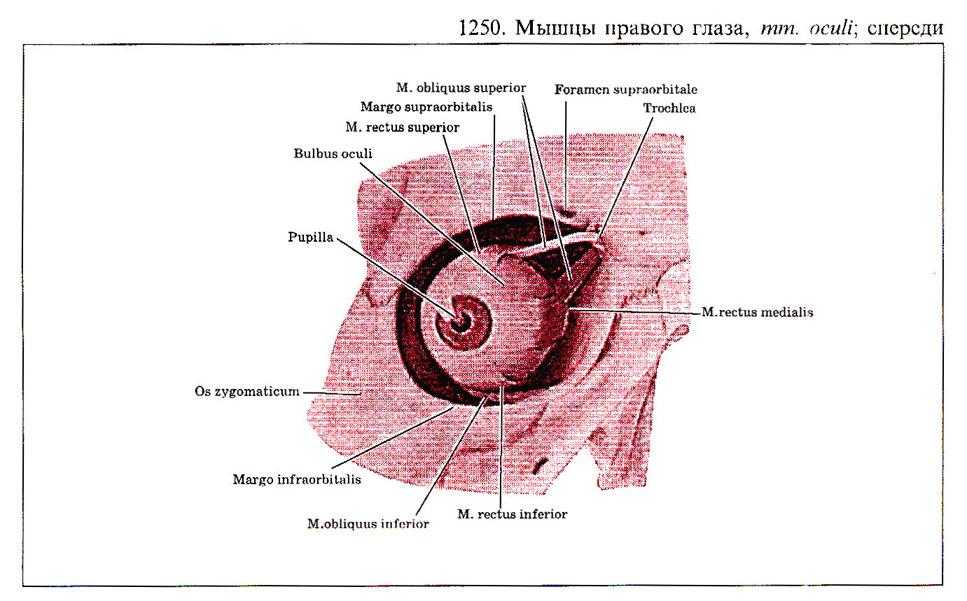
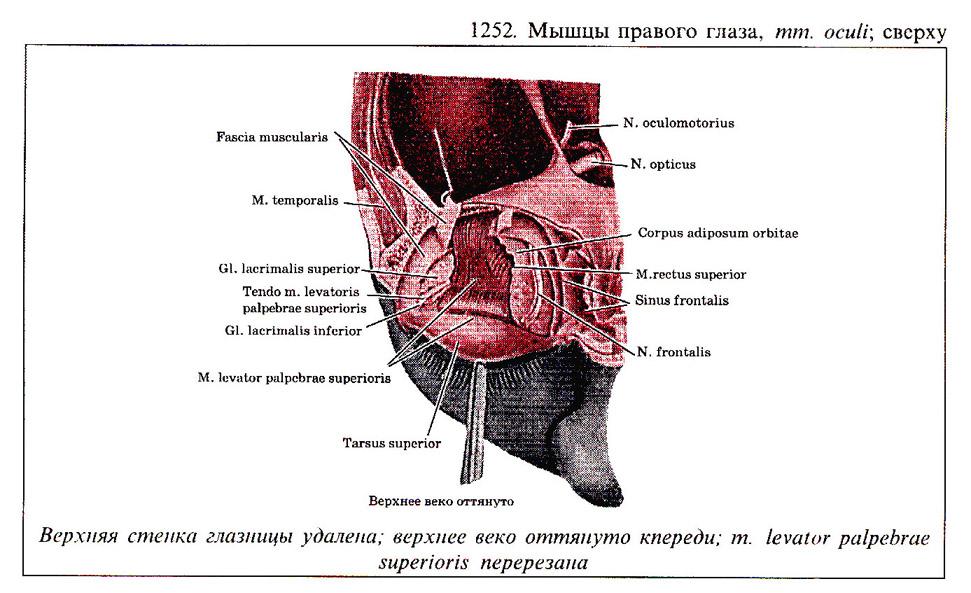
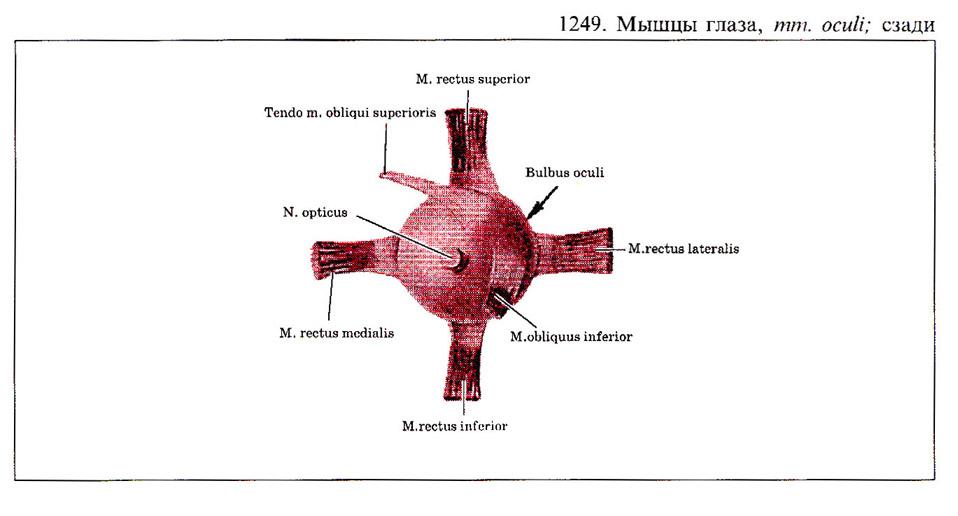
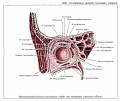
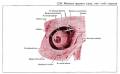
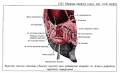
Глаз человека приводится в движение шестью наружными глазными мышцами, которые иннервируются тремя черепными нервами - блоковой нерв (IV) иннервирует верхнюю косую мышцу, отводящий (VI) нерв иннервирует наружную прямую мышцу, а глазодвигательный (III) нерв - четыре остальные наружные мышцы (внутреннюю, нижнюю и верхнюю прямые и нижнюю косую), а также мышцу, поднимающую верхнее веко. В последнее время все большее число работ рассматривает взаимосвязь в управлении быстрыми движениями глаз и процессом моргания (Watanabe et ai.,1980; Voikmann et al., 1982). Не исключено, что процессы подавления зрительного восприятия, сопровождающие акт моргания, играют роль в сохранении стабильного ощущения внешнего мира при движении глаз.
Структура системы иннервации глазодвигательного аппарата весьма сложна. Ядра наружных мышц глаза кошки составляют часть соматической моторной черепномозговой системы. Они располагаются в пределах ретикулярной формации ствола мозга. Ядра VI и IV пары черепномозговых нервов хорошо выделяются и имеют достаточно четкие очертания. Ядро III пары - глазодвигательного нерва состоит из двух отделов - соматического и вегетативного. Главные соматические ядра парные (Oculomotorii lateralis). Добавочные вегетативные парасиматические ядра - непарные (Oculomotorii medialis). Последние представлены ядром Якубовича-Эдингера-Вестфаля и ядром Перля, которое располагается между парой соматических ядер.
Вегетативные ядра участвуют в управлении внутриглазными мышцами. Поражение любого из этих нервов ведет к параличу соответствующих мышц. Наиболее существенным признаком паралича глазных мышц является возникновение двоения при определенном направлении взора (в зависимости от пораженной мышцы). Мотонейроны, к которым относятся перечисленные выше три двигательных черепных нерва, сгруппированы в "ядра" , расположенные в стволе мозга. Степень возбуждения этих нейронов контролируется главным образом нервными клетками "центров контроля взора", связанными с РЕТИКУЛЯРНОЙ ФОРМАЦИЕЙ ВАРОЛИЕВА МОСТА И РЕТИКУЛЯРНОЙ ФОРМАЦИЕЙ СРЕДНЕГО МОЗГА. Характер возбуждения этих нервных клеток определяет характер взаимодействия между шестью наружными мышцами правого и левого глаза.
При нормальном бинокулярном зрении для того, чтобы проекции одного и того же объекта попали в центральные ямки каждого глаза, должен координироваться даже самый незначительный и кратковременный взгляд. На рисунке показана схема работы отдельных глазных мышц. Главное действие - это движение, вызываемое сокращением мышц в том случае, когда глаз находился до этого в нейтральном положении (взгляд вперед). От исходного положения глаза почти не зависят такие движения, которые совершаются наружной прямой (поворот кнаружи или отведение) или внутренней прямой мышцами (поворот кнутри или приведение). Напротив, результат действия всех остальных мышц глаза зависит от положения глазного яблока в глазнице.
Для движения глаз точно по вертикали вверх требуется одновременное сокращение двух мышц - нижней косой и верхней прямой, причем одновременно должно уменьшаться напряжение верхней косой и нижней прямой мышц ( в соответствии с правилами антагонистической иннервации). Наиболее дискуссионной проблемой в физиологии глазодвигательного аппарата оказался вопрос о проприорецепторах глазных мышц. Отсутствие в глазных мышцах обычных лабораторных животных - кролика кошки и собаки - мышечных веретен породило сомнение в наличии вообще каких либо сенсорных окончаний в этих мышцах. Однако в 1946 г. П. Даниель обнаружил в наружных глазных мышцах человека рецептор, очень похожий на веретено. Оказалось, что число веретен весьма значительно, каждая из мышц глаза содержит их около 50.
При общей толщине веретена от 10 до 50 мкм их интрафузальные волокна имеют диаметр 7-20 мкм, а длину от 50 мкм до 1 мм. При столь малых размерах обнаружить эти образования было действительно нелегко. Существенной особенностью глазных веретен является их специфика их дислокации: они встречаются только в дистальном и проксимальном участках мышц и полностью отсутствуют в ее брюшке, т.е. там, где они преимущественно локализованы в мышцах конечностей. По-видимому, веретена в глазных мышцах выступают в роли дублера сухожильных рецепторов - дублера, обладающего к тому же более высокой чувствительностью (что особенно важно для оценки положения глаза в момент фиксационных остановок , может быть, автоматического поддержания фиксации) и сохраняющего в некоторой степени способность к регистрации мышечной динамики. В подтверждение этой мысли можно привести аналогию между способом размещения веретен в мышце глаза и принципом локализации вторичных (статических) эфферентных окончаний в интрафузальных волокнах обычных скелетных мышц. Вторичные окончания локализованы, как правило, по концам волокна, что , по всей вероятности, коррелирует с их функцией статических рецепторов.
Приведенные факты говорят о том, что чувствительность веретен глазной мышцы (и, вероятно, всех ее рецептивных образований) к статическим и динамическим нагрузкам далеко не равноценна. Надо полагать, что эта закономерность обусловлена саккадическим характером движений глаз. В глазодвигательном аппарате тонкая градуальная чувствительность к скорости сокращения теряет свой биологический смысл, т.к. скорость и амплитуда саккады "программируется" заранее и не могут быть изменены во время саккады. В то же время возможность адекватной проприоцептивной оценки позиции глаз в момент фиксации сохраняет свое значение. Очень может быть, что функции глазных веретен человека тесно связаны с поддержанием точных фиксаций при самых различных положениях зрительных осей, т.к. только веретена (в отличие от рецепторов других типов) способны поддерживать - благодаря непрерывным фузимоторным влияниям - постоянную частоту разрядов в работающей мышце.
Афферентное представительство рецепторов наружных мышц глаза в ядерных и надъядерных центрах не вызывает сомнений. Установлено, что импульсы от глазных мышц поступают в двигательные ядра этих мышц, в верхнее двухолмие, в двигательные ядра мышц шеи, и - что особенно важно для дальнейшего анализа - в мозжечковые пути. Одной из главных особенностей наружных мышц глаза является наличие в них тонических волокон (составляющих около 20%). Этот факт был обнаружен на кроликах и описан Д.П.Матюшкиным ( ), а затем подтвержден другими исследователями ( ).Названные волокна обладают основными дефинитивными признаками тонической мускулатуры - они имеют множественные синаптические соединения и сокращаются в результате возникновения местных нераспространяющихся очагов деполяризации. Иными словами, эти волокна не могут быть отнесены к фазическим, а должны быть классифицированы как истинные тонические пучки, сходные с аналогичными пучками низших позвоночных.
Глазные мышцы не единственные обладатели тонических пучков: аналогичные волокна найдены у высших позвоночных и у человека в межреберных мышцах, в диафрагме, в мышцах внутреннего уха. Во всех случаях их функциональное назначение остается неясным. Известно, что глаз человека, несмотря на всю его подвижность, остается в состоянии фиксации 95-97% суммарного времени, а медленные плавные перемещения глаз представляют наряду с саккадами один из главных типов глазодвигательной активности. В то же время данные, приведенные выше, свидетельствуют о том, что тонические мышечные волокна как нельзя лучше приспособлены к поддержанию органов в стабильном положении и плавным градуальным сокращениям. Поэтому естественно было придти к заключению о существовании однозначной морфологически фиксированной корреляции между двумя главными формами глазодвигательной активности (скачками и плавным прослеживанием), с одной стороны, и двумя типами волокон в мышцах глаза (быстрыми фазическими и медленными тоническими ) - с другой.
1.2.БИНОКУЛЯРНАЯ КООРДИНАЦИЯ ДВИЖЕНИЙ ГЛАЗ.
Если рассматривать процесс управления взором (т.е. совместные движения глаз и век при фиксации различных точек пространства), то можно выделить три разные программы:
1.Глаза могут двигаться СОДРУЖЕСТВЕННО верх, вниз вправо или влево, так что их оси остаются параллельными.
2.При смене точек фиксации в пространстве (например, от более близкой к более удаленной) требуются ВЕРГЕНТНЫЕ движения, при которых направление движения одного глаза является примерно зеркальным отражением движения другого. Если фиксируется точка на значительном удалении от наблюдателя, то зрительные оси параллельны. Если взор смещается на более близко расположенный объект, зрительные оси КОНВЕРГИРУЮТ. Для того, чтобы перевести взор на более удаленный предмет, требуется произвести ДИВЕРГЕНЦИЮ, при которой оси глаз несколько разводятся. Если требуется перевести взор с объекта в правой части поля зрения на объект слева, расположенный ближе к наблюдателю, то вергентные и содружественные движения выполняются совместно.
3.ВРАЩАТЕЛЬНЫЕ ДВИЖЕНИЯ ГЛАЗ ВО ФРОНТОПАРАЛЛЕЛЬНОЙ ПЛОСКОСТИ сопровождают наклон головы в сторону. Даже в том случае, когда наклон головы выполняется быстро и на значительный угол, угол поворота глаз не намного превышает 10 градусов.
1.3. ВРЕМЕННЫЕ ХАРАКТЕРИСТИКИ ДВИЖЕНИЙ ГЛАЗ.
По временному критерию движения глаз делит на саккады, периоды фиксации и плавные движения глаз.
САККАДЫ. При свободном рассматривании наши глаза совершают быстрые скачки - саккады - из одной точки фиксации в другую. Амплитуда саккадических движений может варьировать от нескольких угловых минут (при микроскачках) до нескольких градусов (например, если переводить взор из левой части поля зрения в правую). Средняя угловая скорость движения глаз тем выше, чем больше амплитуда саккады и доходит до 200-600 град/сек.
Длительность саккад варьирует от 10 до 80 мс и примерно пропорциональна амплитуде саккады. При осуществлении саккады зрительная система последовательно решает три задачи:
Определяет момент начала скачка
Обеспечивает определенную скорость перемещения глаза
Выбирает момент остановки глаза с таким расчетом, чтобы была достигнута фовеализация новой точки фиксации.
Из перечисленных задач труднее поддается объяснению (в контексте обычных физиологических представлений) третья.
Тезис о запрограммированном механизме саккады основан на том факте, что при движении глаза чувствительность сетчаточного входа резко падает, и поэтому реафферентация по ретинальныму контуру обратной связи существенно затрудняется. Кроме того, скорость скачка превышает скорость, с которой корректировочные ретинальные сигналы передаются по линии обратной связи в глазодвигательные центры. Однако, при ближайшем рассмотрении оказывается, что и принцип программирования саккады, даже возведенный в ранг реально существующего механизма, не может объяснить, почему саккада совершается в определенной точке и как достигается широкое варьирование амплитуды скачков.
ПЕРИОДЫ ФИКСАЦИИ. Даже во время периодов фиксации,которые длятся примерно от 0,15 до 2 сек, глаза немного смещаются за счет глазного тремора, который никогда не прекращается (его амплитуда - несколько угловых минут, а частота находится в пределах 20-150 Гц). Во время продолжительной произвольной фиксации глаз происходит также медленные "дрейф", за счет ко- торого точка фиксации уходит от фиксируемого объекта. С помощью микросаккад происходит компенсация этих дрейфов и восстановление правильной фиксации объекта.
ПЛАВНЫЕ ДВИЖЕНИЯ ГЛАЗ. При необходимости фиксировать движущийся объект требуется выполнение плавных следящих движений глаз. При этом угловая скорость вращательных движений глаз примерно соответствует скорости движения объекта, если скорость последнего не превышает 60-80 град/сек. На плавные следящие движения глаз через переменные интервалы от 0,3 до 2 сек накладываются коррекционные саккады. В этом случае изображение фиксируемой точки предмета остается в пределах 2 град от фовеа. При скорости движения предмета выше 80 град/сек следящие движения глаз происходят значительно медленнее, чем движения предмета, поэтому его изображение перемещается по сетчатке. Если скорость движения предмета не превышает 180 град/сек, то смещение изображения по сетчатке компенсируется с помощью дополнительных саккад большой амплитуды. Координация таких следящих движений глаз обычно осуществляется за счет последующих движений головы. Тогда результирующий сдвиг взора будет (до определенного предела) соответствовать смещению движущегося предмета. Координация движений головы и глаз проводится в центрах управления взора, расположенных в стволе мозга.
3. ФИЛОГЕНЕТИЧЕСКИЕ АСПЕКТЫ ГЛАЗОДВИГАТЕЛЬНОЙ АКТИВНОСТИ
1. На всех этапах эволюции развитие глазодвигательного аппарата тесно коррелирует с развитием зрительной рецепции и образует с ней единое функциональное целое.
2. Филогенез глазодвигательного аппарата (ГДА) не может быть представлен как последовательный переход от примитивных реакций к сложному глазодвигательному поведению: принципы поведенческой целостности и целесообразности характерны для ГДА уже на самых низших этапах его развития, т.е. у низших позвоночных.
3. На отдельных филогенетических срезах в развитии ГДА наблюдается весьма пестрая картина. Это обусловлено тем, что специфика ГДА (как и зрения в целом) подчинена особенностям экологии, которая может сильно различаться в пределах одного класса животных. Поэтому в филогенезе ГДА трудно проследить единую линию развития.
4. Наиболее древней формой ГДА является, по-видимому, компенсаторные повороты глазных яблок, возникающие при поворотах тела животного и направленное на удержание поля зрения. (Если брать ГДА более широко - с учетом работы внутренних глазных мышц, - то придется, вероятно, признать еще большую древность вегетативных зрачковых реакций.)
5. Внешний глазодвигательный аппарат закладывается, по всей вероятности, как единое целое - в совокупности нескольких пар мышц, способствуя адекватной зрительной ориентировке животного в трехмерном пространстве.
6. Главная форма ГДА - плавные компенсаторные повороты и саккады - имеют место у всех позвоночных, причем их динамические характеристики обладают несомненным постоянством на всех этапах эволюции.
7. Степень выраженности ГДА сильно варьирует в связи с экологией животного и непосредственно зависит от широты поля зрения, подвижности головы, уровня фовеализации сетчатки и формы центральной зоны, развития бинокулярности, расположения глаз (фронтального, бокового, косого).
8. Стабильность основных типов ГДА на протяжении всего филогенеза позвоночных свидетельствует о ведущей роли стволовых структур мозга в организации базовых форм движений глаз
9. Управление ГДА осуществляется по иерархическому многоуровневому принципу. По мере развития более высоких (по сравнению со стволом ) мозговых уровней базовые виды ГДА последовательно включаются в новые, более гибкие и совершенные формы поведения.
10. На всех этапах филогенеза глазодвигательный контроль сочетает в себе все ригидные, наследственно фиксированные звенья с пластичными элементами, функционирующими по принципу систем с обратной связью.
11. Управление ГДА изначально имеет полисенсорный характер. Оно обусловлено интеграцией ретинальных, вестибулярных, проприоцептивных, слуховых, тактильных (и, вероятно, других) сигналов, благодаря чему движение глаз оказывается органически включенным в общую моторику и согласуются со "схемой тела".
12. Наиболее поздним приобретением является плавное фовеальное прослеживание движущихся объектов, формирующееся только на уровне приматов и экологически обусловленное наивысшим развитием тонкого бинокулярного зрения в сочетании со зрительно контролируемым манипулированием мелкими предметами.
ПЕРВЫЙ УРОВЕНЬ ГЛАЗОДВИГАТЕЛЬНОЙ СИСТЕМЫ
Первый уровень глазодвигательной системы включает в себя аппарат наружных мышц глаза и аппарат, их иннервирующих. Основой поток управляющих сигналов достигает наружные мышцы по трем парам черепно-мозговых нервов: глазодвигательному, блоковому, отводящему (III,IV,VI). Перекрещивающимся нервом является только VI, остальные - неперекрещивающиеся. Не исключено, что в системе наружных мышц, как во многих других нейронных системах, предусмотрен запасной путь иннервации по ветвям автономной нервной системы. Так, стимуляция цилиарного ганглия может вызвать сокращение наружных мышц глаза, а его разрушение - дегенерацию их волокон. К первому уровню может быть отнесен и нерв V, по которому поступает в мозг основной поток проприоцептивной информации. Например, найдено, что после пресечения V нерва у 6-недельных котят, т.е. в отсутствии проприорецепции, резко уменьшилось (более чем в 6 раз) число ориентационно-избирательных полей у нейронов коры.
ВТОРОЙ УРОВЕНЬ ГЛАЗОДВИГАТЕЛЬНОЙ СИСТЕМЫ
Второй уровень глазодвигательной системы включает в себя ряд "надъядерных" структур, которые являются последней инстанцией, запускающей исполнительные мышечные глазодвигательные механизмы первого уровня системы. К ним относят: ядра ретикулярной формации, структуры моста, некоторые ядра покрышки среднего мозга и ряд других образований , которые могут быть отнесены к центрам управлениям движениями глаз различных направлений (центр вертикального , горизонтального взора, вергентных движений и т.д.).
Следует отметить, что такое гипотетическое представление о существовании отдельных центров взора или центров управления теми или иными движениями глаз имеет противников, которые склонны придерживаться принципа распределенного управления движениями глаз. В последнее время получен ряд фактов, свидетельствующих об участии ядер моста в глазодвигательном акте. Действительно, эта весьма загадочная для физиологов зрения структура тесно связана со многими образованиями зрительно-глазодвигательной системы. На ядра моста проецируются нейроны многих корковых зрительных и ассоциативных полей. На ядра моста у кошки и обезьяны проецируются кроме поля 17 также поля 18,19,20,21 и зоны латеральной супрасильвиевой борозды (Itoh, 1977) и НКТ (Edwards et al.,1974). В свою очередь нейроны тегментального ретикулярного ядра моста посылают аксоны в кору мозжечка. В морфофизиологических экспериментах показано, что paraflocculus и uvula мозжечка получают больше корковых зрительных входов через ядра моста, чем другие церебеллярные области.
Известно, что в свою очередь paraflocculus имеет выраженные проекции на латеральное ядро мозжечка, которое проецируется к различным образованиям ствола мозга, связанным с контролем за движениями глаз - промежуточные и глубокие слои ПБЧ, ядро Даркшевича и интерстициальное ядро Кахаля.
Возможны также прямые связи латерального ядра мозжечка с глазодвигательными ядрами (Carpenter, 1964). В экспериментах установлено, что хотя церебеллоэктомированные животные (обезьяны) могут осуществлять нормальные саккадические движения (Westheimer, Blair,1974), тем не менее у таких обезьян и обезьян с локальными разрушениями различных участков мозжечка наблюдается ряд серьезных нарушений. К ним относятся, например, нарушение процесса продуцирования плавных следящих движений, появление спонтанного нистагма, существенное затруднение удержание отклоненного взора и самого акта фиксации. Однако при этом могут быть также затронуты и саккады, которые в этом случае становятся менее точными. Специфическая роль мозжечка в контроле саккадических движений глаз в свете существующих данных состоит в модулирующих влияниях двух типов на стволовые механизмы генерации саккад:
а) обеспечение точности саккад
б) обеспечение пластичных перестроек (перекалибровка) при разного рода нарушениях или изменениях окружающей среды.
Тонкий нейрофизиологический анализ мозжечковых структур показал, что функции этого органа имеют несомненное отношение к глазодвигательной активности вообще и к организации саккады в частности. На этом основании сложилось аргументированное представление о мозжечке как о "калибраторе" саккады, т.е. такой инстанции, от которой зависит длительность саккады и моменты ее начала и конца. Несколько упрощенно мозжечок может быть представлен как гигантский блок линий задержки саккадического сигнала. Выборочное включение той или иной линии определяется ситуацией, задачей и характером пластических сенсомоторных навыков, которые подвержены постоянными изменениями. При такой трактовке функций мозжечка он выступает как реальная структура, обеспечивающая антиципирующие моторные преднастройки (при саккадах, других формах глазодвигательной активности и общей двигательной активности).
Функциональное предназначение структур второго уровня можно видеть по тем расстройствам глазодвигательного аппарата, которые наступают после поражения надъядерных образований. Это выпадение или нарушение вертикальных, горизонтальных, вергентных движений глаз, парезы или параличи глазных мышц, сопровождающиеся грубыми нарушениями взора. Можно предполагать, что в задачу структур второго уровня входит также функция переключения управляющих движениями глаз сигналов, поступающих от различных сенсорных систем мозга, - зрительной, вестибулярной, слуховой, систем управления движениями головы и тела.
Подобные мультисенсорные влияния испытывают на себе структуры и более высоких уровней зрительно-глазодвигательной системы. Однако мультимодальные посылки (от слуховой, вестибулярной, соматосенсорной систем), поступающие на эти уровни системы, только лишь корректируют процесс формирования выходных сигналов этих уровней, имея право "совещательного голоса", тогда, как сигналы от тех же систем, поступающие на второй уровень, имеют возможность непосредственно управлять движениями глаз.
По-видимому, именно у структур второго уровня существует возможность переключать управляющие воздействия на движения глаз, поступающие от различных сенсорных систем. Определенное подтверждение такому предположению можно видеть в работах, показывающих конвергенцию сигналов от различных сенсорных систем на глазодвигательные структуры второго уровня. Данные об этом можно найти, например, в руководстве Д.Шаде, Д.Форда (1976) или в монографии Р.Карпентера (1977).
ТРЕТИЙ УРОВЕНЬ ГЛАЗОДВИГАТЕЛЬНОЙ СИСТЕМЫ
В отличие от первого и второго этот уровень включает ряд структур, связанных с переработкой информации в прямом пути передачи зрительных сигналов. Согласно рассматриваемой классификации этому уровню принадлежат: базальные ганглии, ПБЧ, мозолистое тело, НКТ (наружные коленчатые тела). К нему же относят и область внутренней капсулы. Электрическое раздражение проходящих в ней аксонов вызывает контрлатеральное движение глаз ( по отношению к раздражаемому полушарию). По-видимому, к этому уровню может быть отнесен комплекс ядер подушки, а также ряд других ядер таламуса, электрическое раздражение которых вызывает определенным образом организованные движения глаз.
Третий уровень по сравнению с более низкими имеет рад функциональных особенностей:
во-первых, в него входят подкорковые структуры зрительного управления движениями глаз и ряд ассоциативных образований, выполняющих сенсорно-интегративные функции в зрительно-глазодвигательной системе управления;
во-вторых, если второй уровень включает в себя центры управления (запуска) отдельными блоками программ движений глаз, центр вертикально-горизонтального взора, конвергенции и т.д., то структуры третьего уровня выполняют задачи координации работы этих центров при выполнении ими отдельных блоков программ движений;
в-третьих, этот уровень участвует в интеграции сигналов, обеспечивающих мультисенсорное управление движениями глаз и координации движений в системе глаз-голова-тело;
в-четвертых, а этом уровне происходит процесс перекодирования результата сенсорного описания внешнего мира на "язык" двигательных программ, выполняемых с помощью двигательных полей нейронов структур этого уровня.
Для успешного выполнения этой задачи структурам третьего уровня необходимо преобразовать различные описания внешнего мира, производимые каждой из сенсорных систем, в некую единую систему описания пространства внешнего мира, в которой функционирует глазодвигательный аппарат.
В настоящее время есть все основания полагать, что подкорковые структуры глазодвигательной системы включают в себя пути управления непроизвольными движениями глаз. В этом аспекте многое определили опыты, позволившие произвести анализ типов движений глаз, вызываемой локальной электрической стимуляцией ряда подкорковых структур, входящих в третий уровень описываемой системы.
Наиболее изучен с помощью методики электрического раздражения феномен вызова движений глаз структурами ПБЧ. Оказалось (Guitton et al.,1980; Roucoux et al., 1980), что локальное раздражение этой структуры может вызвать не только движения глаз различных типов и направлений, но и координированные движения глаз, головы и тела животного. Определены проекционные зоны ПБЧ, раздражения которых сопровождаются движениями либо только глаз животного, либо координированными сочетанными движениями глаз-голова. Движения только глаз вызываются из проекционной зоны ПБЧ, включающей область, окружающую фовеа (area centralis) примерно на 20 угл. град. При помещении раздражающего электрода на большем удалении от проекции фрвеа в ПБЧ вызывались совместные движения глаз и головы или даже (на дальней периферии) и тела животного. По-видимому, результаты этих исследований вскрывают механизм фовеационного рефлекса при его выполнении за счет только либо глазодвигательной системы, либо совместного наведения взора на цель с помощью глаз, головы и тела наблюдателя.
Обнаружение существования проекций из ретикулярной части черной субстанции в верхние бугорки стимулировало целую серию работ, посвященных изучению функциональной значимости недофаминэргичесикх проекций из черной субстанции в организации саккадических движений глаз. Есть данные о свойствах нейронов хвостатого ядра, возможных афферентах, обеспечивающих участие хвостатого ядра в процессе глазодвигательного контроля.
Приведены литературные данные, касающиеся вовлечения нейцроных попукляций комплексов внутренней медулярной полоски (интраоламинарные ядра, медиодорсальное ядро, латеральное дорсальное ядро, вентролатеральное ядро и предние ядра) таламуса (КВМП) в процессе генерации саккадических движений глаз.
Описаны различные типы нейронной активности, предшествующие генерации саккадических движений глаз, коррелирующих с положением глаз в орбите, при спонтанных и зрительно инициируемых саккадических движений глаз. Авторы считают возможным, что генерируемые нейронами КВМП потенциалы инициируют и останавливают обработку и регулируют передачу информации между другими центрами саккадической системы.
ЧЕТВЕРТЫЙ УРОВЕНЬ ГЛАЗОДВИГАТЕЛЬНОЙ СИСТЕМЫ
Этот уровень чаще всего рассматривается в литературе в двух аспектах: клиницистами по картинам нарушений глазодвигательного акта при поражении корковых структур и в опытах на животных с электростимуляцией различных корковых отделов мозга. К этому уровню системы относят: зрительную область коры (17-19 поля), зону Клера-Бишопа, супрасильвиеву, эктосильвиеву области, инсулярную и глазодвигательную кору. Перечисленные структуры имеют проекционные связи с ПБЧ; частично с нейронами дорсального и вентрального НКТ (наружных коленчатых тел), подушкой, ретикулярным ядром таламуса.
Все эти структуры так или иначе участвуют в организации глазодвигательного акта. Из всех уровней глазодвигательной системы наименее изучен корковый уровень зрительного управления движениями глаз. До сих пор используется сформулированные в работах Холмса (Holmes,1938) представления о "заднем" - затылочном и "переднем" (премоторная кора) корковых центров управления движениями глаз. Исследования этих зон управления взором было начато задолго до работ Холмса еще в середине прошлого века. Наибольший вклад в их изучение внесли исследования В.М. Бехтерева (Бехтерев, 1907), работы А.Феррьера, Е. Гитцига, Г. Фритша (Ferrier,1876; Fritsch, Hitzig, 1870) и других исследователей. Было показано, что передний центр, в отличие от затылочного - зрительного, участвует в организации произвольных движений глаз, тогда как затылочный - организует непроизвольные движения. Затылочный и лобный центры в какой-то степени автономны и являются реципрокно взаимодействующими подсистемами управления движениями глаз.
Опыты с электрическим раздражением этих структур выявили определенное функциональное доминирование лобного центра над затылочным. Это заключение сделано на основании сравнительно большей чувствительности структур моторной зоны к электрическому раздражению и меньшему латентному периоду движений, вызываемых стимуляцией переднего центра (Brown, Graham,1927).Естественно, что эта автономность весьма условна, т.к. большая часть необходимой для организации движений глаз зрительной информации поставляется в моторную зону не минуя затылочные зрительные поля коры.
К тому же по результатам опытов с электрическим раздражением отдельных структур трудно судить о реальных временных взаимоотношениях работы этих структур при их функционировании в реальных условиях видения. Несомненно, что во многом условно и само деление на передний и задний глазодвигательные центры. Движения, организуемые задним центром, относятся к непроизвольным, или автоматическим. Передний центр, в отличие от заднего, участвуя в организации произвольных, многоходовых движений глаз, оперирует с управляющими сигналами, порождаемыми на основании значительно большего объема информации, используя такие, пока структурно не формализуемые "факторы", как, например, память, внимание и т.д.
Взаимосвязь зрительных ощущений и движений глаз опосредована разнообразными связями кортикальных зрительных полей и мотонейронов глазных мышц, расположенных в стволе головного мозга. Область V2, например, проецируется в Colliculi superiores, от которой аксоны глазодвигательных нейронов поднимаются в парамедманную ретикулярную формацию варолиева моста и ретикулярную формацию среднего мозга. Из экстрастриарных зрительных полей есть проекции в префронтальные регионы коры БП, которые служат самопроизвольному управлению движениями глаз (область 8) и координации движений "рука-рот". Из этих регионов, как и их области 7 париетальной коры БП, есть проекции в центры управления взором ствола мозга и назад, в окципитальные зрительные поля коры БП. Для афферентных проекций специфических движущихся зрительных сигналов путь от сетчатки в верхние бугорки четверохолмия имеет важное значение.
Нейроны верхнего двухолмия имеют связи с подушкой таламуса, откуда сигналы поступают в экстрастриарные зрительные пола БП и в область теменной доли. Эти связи обеспечивают также интеграцию движений глаз и зрительных ощущений.
ГЛАЗОДВИГАТЕЛЬНАЯ ПЕТЛЯ. Эту петля можно вполне определенно выделить анатомически, она предназначена для контроля движений глаз. Она начинается от кортикальных полей, которые управляют моторикой глаз, а именно: от фронтального зрительного поля (поле 8 по Бродману) и каудальной частью поля 7 париетальной коры. Поток информации проходит через N.caudatus и дорсомедиальный сектор внутренней части бледного шара или через вентролатеральную часть Pars reticulata черной субстанции. Следующий пункт переключения таламическими ядрами, их которых информация вновь возвращается во фронтальные зрительные поля. Следует упомянуть, что аксоны ретикулярных нейронов дают коллатераль в верхние бугорки четверохолмия, которые, как известно, тоже участвуют в организации движений глаз.
Рисунок показывает позитивную корреляцию между активностью ретикулярных клеток и глазными саккадами. Интенсивность разряда снижается с фазическим течением перед саккадой ,как проявление угнетающего влияния Striatum nigra. Вследствие этого в таламусе или в верхних бугорках возникает фазическая активность. Строгое пространственное разграничение скелетомоторной и глазодвигательной петли проявляется , например, в том, что активность нейронов Pars reticulata коррелирует либо с движениями глаз, либо с орофациальными движениями, но не с обоими видами движений. Активность клеток черной субстанции, Pars reticulata (обезьяны), функциональное состояние которых коррелирует с глазными саккадами. Вертикальным штрихом обозначена отметка времени вспышки света. Обезьяна натренирована переводить взор на новую точку. Примерно через 100 мс после действия светового раздражителя и примерно за 200 мс до начала саккады активность клеток почти полностью подавляется.
4. ИЕРАРХИЧЕСКОЕ УПРАВЛЕНИЕ ДВИЖЕНИЯМИ ГЛАЗ
Сравнительный анализ нервно-мышечной организации глазодвигательной активности и других соматических движений позволяет выявить черты как сходства, так и р
Похожие статьи
Добавь в закладки
|